


- 162.50 KB
- 18页

- 1、本文档共5页,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,可选择认领,认领后既往收益都归您。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细先通过免费阅读内容等途径辨别内容交易风险。如存在严重挂羊头卖狗肉之情形,可联系本站下载客服投诉处理。
- 文档侵权举报电话:19940600175。
18第一章糖一、糖的概念糖类物质是多羟基(2个或以上)的醛类(aldehyde)或酮类(Ketone)化合物,以及它们的衍生物或聚合物。据此可分为醛糖(aldose)和酮糖(ketose)。还可根据碳层子数分为丙糖(triose),丁糖(terose),戊糖(pentose)、己糖(hexose)。最简单的糖类就是丙糖(甘油醛和二羟丙酮)由于绝大多数的糖类化合物都可以用通式Cn(H2O)n表示,所以过去人们一直认为糖类是碳与水的化合物,称为碳水化合物。现在已经这种称呼并恰当,只是沿用已久,仍有许多人称之为碳水化合物。二、糖的种类根据糖的结构单元数目多少分为:(1)单糖:不能被水解称更小分子的糖。(2)寡糖:2-6个单糖分子脱水缩合而成,以双糖最为普遍,意义也较大。(3)多糖:均一性多糖:淀粉、糖原、纤维素、半纤维素、几丁质(壳多糖)不均一性多糖:糖胺多糖类(透明质酸、硫酸软骨素、硫酸皮肤素等)(4)结合糖(复合糖,糖缀合物,glycoconjugate):糖脂、糖蛋白(蛋白聚糖)、糖-核苷酸等(5)糖的衍生物:糖醇、糖酸、糖胺、糖苷三、糖类的生物学功能(1)提供能量。植物的淀粉和动物的糖原都是能量的储存形式。(2)物质代谢的碳骨架,为蛋白质、核酸、脂类的合成提供碳骨架。(3)细胞的骨架。纤维素、半纤维素、木质素是植物细胞壁的主要成分,肽聚糖是细胞壁的主要成分。(4)细胞间识别和生物分子间的识别。细胞膜表面糖蛋白的寡糖链参与细胞间的识别。一些细胞的细胞膜表面含有糖分子或寡糖链,构成细胞的天线,参与细胞通信。
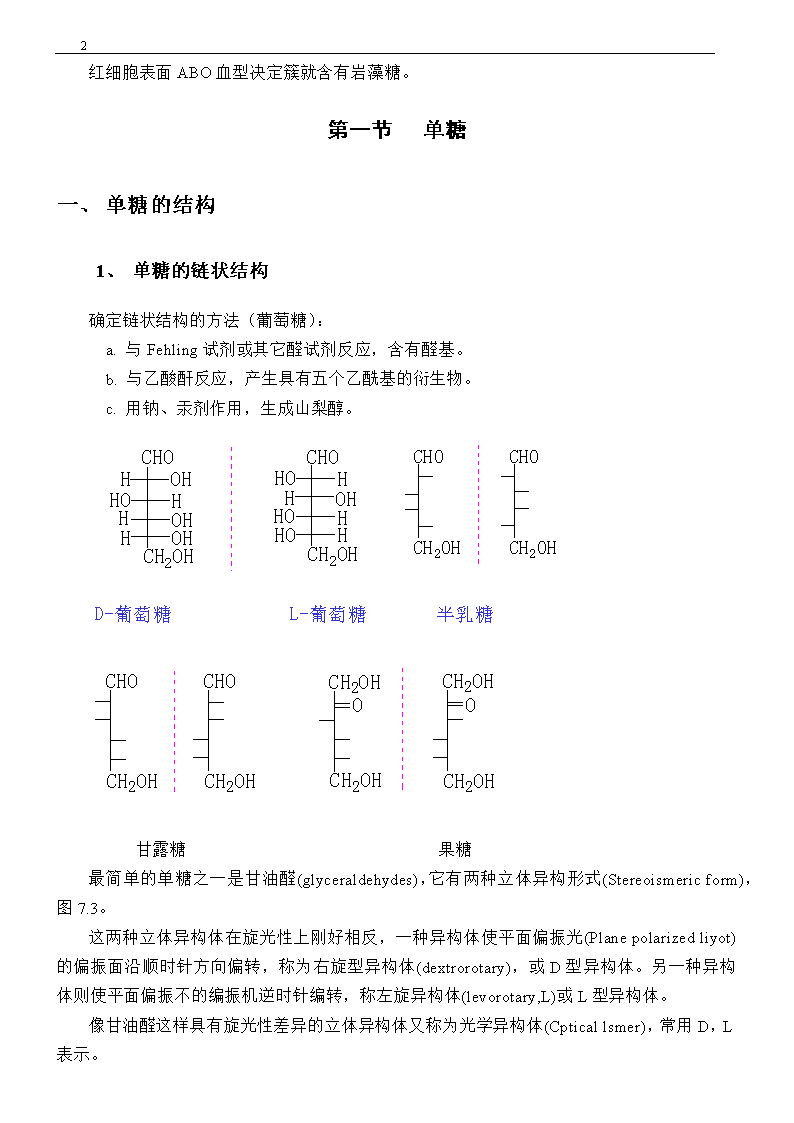
18红细胞表面ABO血型决定簇就含有岩藻糖。第一节单糖一、单糖的结构1、单糖的链状结构确定链状结构的方法(葡萄糖):a.与Fehling试剂或其它醛试剂反应,含有醛基。b.与乙酸酐反应,产生具有五个乙酰基的衍生物。c.用钠、汞剂作用,生成山梨醇。D-葡萄糖L-葡萄糖半乳糖甘露糖果糖最简单的单糖之一是甘油醛(glyceraldehydes),它有两种立体异构形式(Stereoismericform),图7.3。这两种立体异构体在旋光性上刚好相反,一种异构体使平面偏振光(Planepolarizedliyot)的偏振面沿顺时针方向偏转,称为右旋型异构体(dextrorotary),或D型异构体。另一种异构体则使平面偏振不的编振机逆时针编转,称左旋异构体(levorotary,L)或L型异构体。像甘油醛这样具有旋光性差异的立体异构体又称为光学异构体(Cpticallsmer),常用D,L表示。
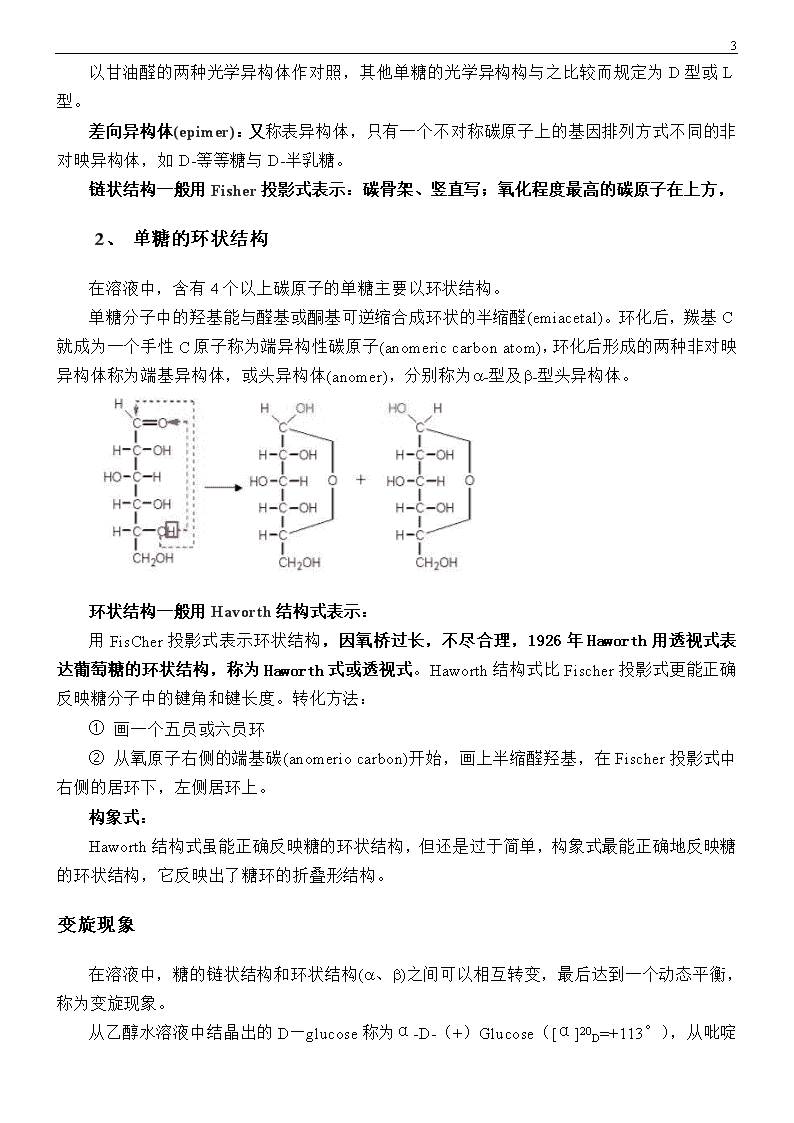
18以甘油醛的两种光学异构体作对照,其他单糖的光学异构构与之比较而规定为D型或L型。差向异构体(epimer):又称表异构体,只有一个不对称碳原子上的基因排列方式不同的非对映异构体,如D-等等糖与D-半乳糖。链状结构一般用Fisher投影式表示:碳骨架、竖直写;氧化程度最高的碳原子在上方,1、单糖的环状结构在溶液中,含有4个以上碳原子的单糖主要以环状结构。单糖分子中的羟基能与醛基或酮基可逆缩合成环状的半缩醛(emiacetal)。环化后,羰基C就成为一个手性C原子称为端异构性碳原子(anomericcarbonatom),环化后形成的两种非对映异构体称为端基异构体,或头异构体(anomer),分别称为a-型及b-型头异构体。环状结构一般用Havorth结构式表示:用FisCher投影式表示环状结构,因氧桥过长,不尽合理,1926年Haworth用透视式表达葡萄糖的环状结构,称为Haworth式或透视式。Haworth结构式比Fischer投影式更能正确反映糖分子中的键角和键长度。转化方法:①画一个五员或六员环②从氧原子右侧的端基碳(anomeriocarbon)开始,画上半缩醛羟基,在Fischer投影式中右侧的居环下,左侧居环上。构象式:Haworth结构式虽能正确反映糖的环状结构,但还是过于简单,构象式最能正确地反映糖的环状结构,它反映出了糖环的折叠形结构。变旋现象在溶液中,糖的链状结构和环状结构(a、b)之间可以相互转变,最后达到一个动态平衡,称为变旋现象。从乙醇水溶液中结晶出的D—glucose称为α-D-(+)Glucose([α]20D=+113°
18),从吡啶溶液中结晶出的D—glucose称为β-D-(+)glucose([α]20D=+18.7°)。将a-D-(+)葡萄糖与b-D-(+)葡萄糖分别溶于水中,放置一段时间后,其旋光率都逐渐转变为+52.7°C。原因就是葡萄糖的不同结构形式相互转变,最后,各种结构形式达到一定的平衡,其中a型占36%,b型占63%,链式占1%。1、构型与构象构型:分子中由于各原子或基团间特有的固定的空间排列方式不同而使它呈现出不同的较定的立体结构,如D-甘油醛与L-甘油醛,D-葡萄糖和L葡萄糖是链状葡萄糖的两种构型,a-D-葡萄糖和b-D-葡萄糖是环状葡萄糖的两种构型。一般情况下,构型都比较稳定,一种构型转变另一种构型则要求共价键的断裂、原子(基团)间的重排和新共价键的重新形成。构象:由于分子中的某个原子(基团)绕C-C单键自由旋转而形成的不同的暂时性的易变的空间结构形式,不同的构象之间可以相互转变,在各种构象形式中,势能最低、最稳定的构象是优势对象。2、构型与旋光性旋光性是分子中具有不对称结构的物质的一种物理性质。显然,构型不同旋光性就不同。构型是人为规定的,旋光性是实验测出的。因此,构型与旋光性之间没有必然的对应规律,每一种物质的旋光性只能通过实验来确定。二、单糖的物理化学性质(一)物理性质旋光性:是鉴定糖的一个重要指标糖都有使平面偏振光发生偏转的能力,即糖的旋光性,是因为糖都具有手性碳。糖的旋光性和旋光度由糖分子中的所有手性碳上的羟基方向所决定。糖的旋光性以右旋(以d或+表示)或左旋(以l或-表示)。旋光性与D-或L-构型没有必然联系。但同一化合物的D-型和L-型旋光方向相反。在一定条件下,测定一定浓度糖溶液的旋光度,可以计算共比旋光度
18甜度:以蔗糖的甜度为标准糖甜度糖甜度果糖173.3鼠李糖32.5转化糖130麦芽糖32.5蔗糖100半乳糖32.1葡萄糖74.3棉子糖22.6木糖40乳糖16.1溶解性:单糖分子,甘油醛外,有许多亲水基团,易溶于水,不溶于乙醚、丙酮等有机溶剂。(一)化学性质1、异构化2、氧化反应(1)土伦试剂、费林试剂氧化(碱性氧化)醛糖与酮糖都能被象土伦试剂或费林试剂这样的弱氧化剂氧化,前者产生银镜,后者生成氧化亚铜的砖红色沉淀,糖分子的醛基被氧化
18为羧基。凡是能被上述弱氧化剂氧化的糖,都称为还原糖,所以,果糖也是还原糖。果糖具有还原性的原因:差向异构化作用——果糖在稀碱溶液中可发生酮式-烯醇式互变,酮基不断地变成醛基(土伦试剂和费林试剂都是碱性试剂,故酮糖能被这两种试剂氧化)。(2)溴水氧化(酸性氧化)溴水能氧化醛糖,但不能氧化酮糖,因为酸性条件下,不会引起糖分子的异构化作用。可用此反应来区别醛糖和酮糖。(3)硝酸氧化稀硝酸的氧化作用比溴水强,能使醛糖氧化成糖二酸。(4)高碘酸氧化糖类象其他有两个或更多的在相邻的碳原子上有羟基或羰基的化合物一样,也能被高碘酸所氧化,碳碳键发生断裂。反应是定量的,每破裂一个碳碳键消耗一摩尔高碘酸。因此,此反应是研究糖类结构的重要手段之一。单糖氧化形成的羟基可以进一步形成环状内酯(Lactone)。内酯在自然界中很普遍,如L-抗坏血酸(L-ascorbioacid),又称VC(Vitamcnc),就是D-葡萄糖酸的内酯衍生物。分子量176.1,它在体内是一种强还原剂。豚鼠(guineapig)、猿(ape)和人不能合成Vc,从能合成Vc的肝脏微粒体中分离到合成Vc的三种酶,人和猿缺乏gulonolactoneoxidase)。缺乏抗坏血酸将导致坏血病(scurvy),龄龈(gum)、腿部等开始出血,肿胀,逐渐扩展到全身,柑橘类果实(citrusfrait)中含有丰富的Vc。1、还原反应单糖可以被还原成相应的糖醇(Sugaralcohol)。D-葡萄糖被还原成D-葡萄糖醇,又称山犁醇(D-Sorbitol)。甘露糖产生甘露醇,核糖产生核醇。糖醇主要用于食品加工业和医药,山犁醇添加到糖果中能延长糖果的货架期,因为它能防止糖果失水。用糖精处理的果汁中一般都有后味,添加山犁醇后能去除后味。人体食用后,山犁醇在肝中又会转化为果糖。
181、酯化生物体中最常见也是最重要的糖酯是磷酸糖酯和硫酸糖酯。磷酸糖酯及其衍生物是糖的代谢活性形式(糖代谢的中间产物)。硫酸糖酯主要发现于结缔组织的蛋白聚糖中(Proteoglycan),由于硫酸糖酯带电荷,因此它能结合大量的水和阳离子。葡萄糖的核苷二磷酸酯,如UDPG参与多糖的生物合成。2、糖苷化糖分子中的活泼半缩醛羟基与其它含羟基的化合物(如醇、酚),含氮杂环化合物作用,失水而生成缩醛的反应称为成苷反应。其产物称为配糖物,简称为“苷”,全名为某糖某苷。注意几点:①苷似醚不是醚,它比一般的醚键易形成,也易水解。②苷用酶水解时有选择性③糖苷没有变旋光现象,没有还原糖的反应。④糖苷在自然界的分布极广,与人类的生命和生活密切相关。3、糖脎反应(亲核加成)糖脎反应发生在醛糖和酮糖的链状结构上。糖脎易结晶,可以根据结晶的形状,判断单糖的种类。(过量)
187、强酸催化脱水作用单糖在强酸作用下,受热脱水生成糠醛或糠醛衍生物。糠醛是多种化工合成所需的原料。糠醛或羟甲基糠醛能与某些酚类作用生成有色的缩合物,利用这一性质,可进行糖的定性定量测定。一、重要的单糖(1)丙糖:D-甘油醛二羟丙酮(2)丁糖:D-赤鲜糖D-赤鲜酮糖(3)戊糖:D-核糖D-脱氧核糖D-核酮糖D-木糖D-木酮糖核糖与脱氧核糖(4)己糖:D-葡萄糖(a-型及b型)D-果糖(5)庚糖:D-景天庚酮糖7-磷酸酯存在于磷酸无糖途径
18一、重要的单糖衍生物1、糖醇木糖醇;肌醇(环六己醇)(肌肌醇);肌醇-1,4,5三磷酸(IP3);植酸2、糖醛酸单糖的伯醇基被氧化成-COOH。动物体内有两种很重要的糖醛酸:a-D-葡萄醛酸和差向异构物b-L-艾杜糖醛酸,它们在结缔组织中含量很高。葡萄糖醛酸是肝脏内的一种解毒剂,它与类固醇、一些药物、胆红素(血红蛋白的降解物)结合增强其水溶性,使之更易排出体外。3、氨基糖(糖胺,aminosugar,glycosamine)单糖的一个羟基(通常是C2位)被氨基取代。常见的氨基糖有D-葡萄糖胺(D-glucosamine)和D-半乳糖胺(D-galactosamine)。氨基糖的氨基还经常被乙酰化形成N-乙酰糖胺、N-乙酰胞璧酸等4、糖苷单糖的半缩醛羟基与其它分子的醇、酚等羟基缩合,脱水生成缩醛式衍生物,称糖苷Glycoside。半缩醛部分是Glc,称Glc糖苷。半缩醛部分是Gal,称Gal糖苷。O糖苷、N糖苷、S糖苷。糖苷物质与糖类的区别:糖是半缩醛,不稳定,有变旋;苷是缩醛,较稳定,无变旋。糖苷大多数有毒。5、脱氧糖重要的有6-脱氧D-甘露糖,L-岩藻糖(L-fucose)和2-脱氧D-核糖。岩藻糖常见于一些糖蛋白中,如红细胞表面ABO血型决定簇。第二节双糖和三糖
18双糖在自然界中含量也很丰富,它是人类饮食中主要的热源之一。在小肠中,双糖必须在酶的作用下水解成单糖才能被人体吸收。如果这些酶有缺陷的话,那么人体摄入双糖后由于不能消化它就会出现消化病。未消化的双糖进入大肠,在渗透压的作用下从周围组织夺取水分(腹泻,diarrhea),结肠中的细菌消化双糖(发酵)产生气体(气胀和绞痛或痉孪)。最常见的双糖消化缺陷是乳糖过敏,就是由于缺乏乳糖酶(Lactose),解决办法就是乳糖酶处理食物或避免摄入乳糖。一、麦芽糖(maltose,maltsugar)它是直链淀粉的水解中间物(a-麦芽糖),在自然界中似乎并不存在天然的麦芽糖。结构:两分子a-葡萄糖,a(1-4)糖苷键。a-麦芽糖(葡萄糖-a,a(1-4)-葡萄糖苷)b-麦芽糖[葡萄糖-a,b(1-4)-葡萄糖苷]性质:①变旋现象,在水溶解中形成a、b和开链的混合物②具有还原性③能成脎异麦芽糖:a(1-6)键型,支链淀粉和糖元的水解产物二、蔗糖植物的茎、叶都可以产生蔗糖,它可以在整个植物体中进行运输,也是光合产物的运输形式之一。结构:a-葡萄糖,b-果糖a,b(1-2)糖苷键,无异构体性质:①无变旋现象②无还原性③不能成脎三、乳糖顾名思义,主要存在于哺乳动物的乳汁中结构:b-半乳糖b(1-4)糖苷键a(或b)-葡萄糖。两种异构体。a-Lactose[半乳糖-b,a(1-4)-葡萄糖苷]b-lactose[半乳糖-b,b(1-4)-葡萄糖苷]性质:①有变旋现象②具有还原性③能成脎四、纤维二糖(cellobiose)纤维素的降解产物和基基本结构单位,自然界中不存在游离的纤维二糖结构:两分子b-葡萄糖b-(1,4)糖苷键
18纤维二糖[葡萄糖-b(1,4)-葡萄糖苷]性质:①具有变旋现象②具有还原性③能成脎一、海藻糖两分子α-D-Glc,在C1上的两个半缩醛羟基之间脱水,由α-1.1糖苷键构成。二、棉子糖(三糖)P31结构非还原性三糖第二节寡糖寡糖是指含有2-10个单糖单元的糖类。它们常常与蛋白质或脂类共价结合,以糖蛋白或糖脂的形式存在。连接它们的共价键类型主要两大类:N-糖甘键型和O-糖苷键型。①N-糖苷键型:寡糖链与多肽上的Asn的氨基相连。这类寡糖链有三种主要类型:高甘露糖型,杂合型和复杂型。②O-糖苷键型,寡糖链与多肽链上的Ser或Thr的羟基相连,或与膜脂的羟基相连。第三节多糖多糖是由多个单糖分子缩合脱水而形成的。由于构成它的单糖的种类、数量以及连接方式的不同,多糖的结构极其复杂而且数量、种类庞大。多糖是重要的能量贮存形式(如淀粉和糖原等)和细胞的骨架物质(如植物的纤维素和动物的几丁质),此外多糖还有更复杂的生理功能(如粘多糖和血型物质等)。大部分的多糖类物质没有固定的分子量。多糖的大小从一定程度上可以反映细胞的代谢状态。例如:当血糖水平高时(如饭后),肝脏就合成糖原(glycogen)这时就分子量可达2´107,当血糖水平下降时,肝脏中的酶类就水解糖原,把葡萄糖释放到血液中。多糖在水溶液中只形成胶体,虽然具有旋光性,但无变旋现象,也无还原性。多糖可以分为均一性多糖(由同一种单糖分子组成)和不均一性多糖(由两种或两种以上单糖分子组成)一、均一性多糖
18自然界中最丰富的均一性多糖是淀粉和糖原、纤维素。它们都是由葡萄糖组成。淀粉和糖原分别是植物和动物中葡萄糖的贮存形式,纤维素是植物细胞主要的结构组分。1、淀粉植物营养物质的一种贮存形式,也是植物性食物中重要的营养成分。①直链淀粉许多a-葡萄糖以a(1-4)糖苷键依次相连成长而不分开的葡萄糖多聚物。典型情况下由数千个葡萄糖线基组成,分子量从150000到600000。结构:长而紧密的螺旋管形。这种紧实的结构是与其贮藏功能相适应的。遇碘显兰色图7.30②支链淀粉在直链的基础上每隔20-25个葡萄糖残基就形成一个a-(1-6)支链。不能形成螺旋管,遇碘显紫色。淀粉酶:内切淀粉酶(α-淀粉酶)水解α-1.4键,外切淀粉酶(β-淀粉酶)α-1.4,脱支酶α-1.62、糖元与支链淀粉类似,只是分支程度更高,分支更,每隔4个葡萄糖残基便有一个分支。结构更紧密,更适应其贮藏功能,这是动物将其作为能量贮藏形式的一个重要原因,另一个原因是它含有大量的非原性端,可以被迅速动员水解。糖元遇碘显红褐色。3、纤维素结构:许多b-D-葡萄糖分子以b-(1-4)糖苷键相连而成直链。纤维素是植物细胞壁的主要结构成份,占植物体总重量的1/3左右,也是自然界最丰富的有机物,地球上每年约生产1011吨纤维素,经济价值:木材、纸张、纤维、棉花、亚麻。完整的细胞壁是以纤维素为主,并粘连有半纤维素、果胶和木质素。约40条纤维素链相互间以氢键相连成纤维细丝,无数纤维细丝构成细胞壁完整的纤维骨架。图7.33
18降解纤维素的纤维素主要存在于微生物中,一些反刍动物可以利用其消化道内的微生物消化纤维素,产生的葡萄糖供自身和微生物共同利用。虽大多数的动物(包括人)不能消化纤维素,但是含有纤维素的食物对于健康是必需的和有益的。1、几丁质(壳多糖):N-乙酰-b-D-葡萄糖胺以b(1,4)糖苷链相连成的直链。2、菊糖inulin多聚果糖,存在于菊科植物根部。3、琼脂Ager多聚半乳糖,是某些海藻所含的多糖,人和微生物不能消化琼脂。几种均一多糖的结构、性质比较。P35表1-6二、不均一性多糖不均一性多糖种类繁多。有一些不均一性多糖由含糖胺的重复双糖系列组成,称为糖胺聚糖(glyeosaminoglycans,GAGs),又称粘多糖。(mucopolysaceharides)、氨基多糖等。糖胺聚糖是蛋白聚糖的主要组分,按重复双糖单位的不同,糖胺聚糖有五类:1、透明质酸2、硫酸软骨素3、硫酸皮肤素4、硫酸用层酸5、肝素6、硫酸乙酰肝素第二节结合糖(glycoconjugate)糖与非糖物质共价结合形成的复合物称结合糖(复合糖,糖缀合物),包括糖脂(glycolipids),糖蛋白与蛋白聚糖、肽聚糖(peptidoglycan),糖—核酸。
18一、糖蛋白糖蛋白是由短的寡糖链与蛋白质共价相连构成的分子。其总体性质更接近蛋白质。糖与蛋白质之间以蛋白质为主,其一定部位上以共价健与若干短的寡糖链相连,这些寡糖链常常是具分支的杂糖链,不呈现重复的双糖系列,一般由2-10个单体(少于15)组成,未端成员常常是唾液酸或L-岩藻糖。(一)组成β-D-葡萄糖(Glc)α-D-甘露糖(Man)α-D-半乳糖(Gal)α-D-木糖(Xyl)α-D-阿拉伯糖(Ara)α-L-岩藻糖(Fuc)葡萄糖醛酸(GlcuA)艾杜糖醛酸(IduA)N-乙酰葡萄糖胺(GlcNAG)N-乙酰半乳糖胺(GalNAC)N-乙酰神经氨酸(NeuNAC)即唾液酸(Sia)(二)糖链与蛋白的连接方式糖蛋白的糖肽连接键,简称糖肽键。糖肽链的类型可以概况为:①N-糖苷键型:寡糖链(GlcNAC的β-羟基)与Asn的酰胺基、N-未端的a-氨基、Lys或Arg的W-氨基相连图15②O-糖苷键型:寡糖链(GalNAC的α-羟基)与Ser、Thr和羟基赖氨酸、羟脯氨酸的羟基相连。图16③S-糖苷键型:以半胱氨酸为连接点的糖肽键。④酯糖苷键型:以天冬氨酸、谷氨酸的游离羧基为连接点。(三)糖蛋白中糖链的结构糖蛋白中的糖链变化较大,含有丰富的结构信息。寡糖链往往是受体、酶类的识别位点。1、N-糖苷键型(N-连接)N-糖苷键型主要有三类寡糖链:①高甘露糖型,由GlcNAc和甘露糖组成;②复合型:除了GlcNAc和甘露糖外、还有果糖、半乳糖、唾液酸;③杂合型,包含①和②的特征。图17五糖核心A.高甘露糖型
18中国地仓鼠卵细胞膜图18B.N-乙酰半乳糖型图19C.混合型卵白蛋白的一种糖链图201、O-糖苷键型(O-连接)没有五糖核心。图21人血纤维蛋白溶酶原:图22人免疫球蛋白IgA:(二)糖蛋白的生物学功能(1)糖蛋白携带某些蛋白质代谢去向的信息糖蛋白寡糖链末端的唾液酸残基,决定着某种蛋白质是否在血流中存在或被肝脏除去的信息。A脊椎动物血液中的铜蓝蛋白肝细胞能降解丢失了唾液酸的铜蓝蛋白,唾液酸的消除可能是体内“老”蛋白的标记方式之一。B.红细胞新生的红细胞膜上唾液酸的含量远高于成熟的红细胞膜。用唾液酸酶处理新生的红细胞,回注机体,几小时后全部消失。而末用酶处理的红细胞,回注后,几天以后,仍能在体内正常存活。(2)寡糖链在细胞识别、信号传递中起关键作用淋巴细胞正常情况应归巢到脾脏,而切去唾液酸后,结果竞归巢到了肝脏。
18在原核中表达的真核基因,无法糖基化。糖蛋白可以是胞溶性的,也可以是膜结合型的,可以存在于细胞内在也可存在于细胞间质中。糖蛋白在动植物中较为典型,脊柱动物中糖蛋白尤为丰富,金属转运蛋白(转铁蛋白)、血铜蓝蛋白,凝血因子、补体系统、一些激素,促卵泡素(Follicle-stimulatinghormone,FSH,前脑下垂体分泌,促进卵子和精子的发育)、RNase、膜结合蛋白(如动物细胞膜的Na+-K+-ATPase)、主要组织相容性抗原(majorhistocompatibilityantigen,细胞表面上介导供体器官与受体器官交叉匹配的标识)。绝大多数糖蛋白的寡糖是糖蛋白的功能中心。有些糖蛋白的糖对于糖蛋白自身成机体起着保护作用或润滑作用,如牛的RNaseB(糖蛋白)对热的抗性大于RNaseA,大量的唾液酸能增强唾液粘蛋白的粘性从而增强唾液的润滑性。南极鱼抗冻蛋白的糖组分能与水形氢键,阻止冰品的形成从而提高了抗冻性。糖蛋白在细胞间信号传递方面着更为复杂的作用。Hiv的靶细胞结合蛋白GP120是一个糖蛋白,能与人类靶细胞表面的CD4受体结合从而附着在靶细胞表面,如果去掉GP120的糖部分则不能与CD4受体结合从而失去感染能力。细胞表面的糖蛋白形成细胞的糖萼(糖衣)、参与细胞的粘连,这在胚和组织的生长、发育以及分化中起着关键性作用。一、蛋白聚糖(oroteoglycans)由糖胺聚糖与多肽链共价相连构成的分子,总体性质与多糖更为接近。糖胺聚糖链长而不分支,呈现重复双糖系列结构,其一定部位上与若干肽链相连。由于糖胺聚糖具有粘稠性,所以蛋白聚白又称为粘蛋白、粘多糖–蛋白质复合物等。(一)蛋白聚糖中的糖肽键在蛋白聚糖中已知有三种不同类型的糖肽键:1、D-木糖与Ser羟基之间形成的O-糖肽键;硫酸软骨素硫酸皮肤素硫酸类肝GlcUAβ1→3Galβ1→3Galβ1→4Xyl1→Ser肝素
181、N-乙酰半乳糖胺与Thr或Ser羟基之间形成的O-糖肽键。骨骼硫酸角质素→GalNAcl→6GalNAc→ser(Thr)Sia2→3Gal1→3↗2、N-乙酰葡萄糖胺与Asn之间形成的N-糖肽键;角膜硫酸角质素→GlcNAc—N—Asn.(二)糖白聚糖的生物学功能糖白聚糖主要存在于软骨、键等结缔组织和各种腺体分泌的粘液中,有构成组织间质、润滑剂、防护剂等多方面的作用。二、肽聚糖peptidoglycan是细菌细胞壁的主要成分,草兰氏阳性细菌胞壁所含的肽聚糖占干重的50-80%,草兰氏阴性细菌胞壁所含的肽聚糖占干重的1-10%糖链由N-乙酰葡萄糖胺和N-乙酰胞壁酸通过β-1.4糖苷键连接而成,糖链间由肽链交联,构成稳定的网状结构,肽链长短视细菌种类不同而异。图14组成及结构特点(金黄色葡萄球菌)1.G—M聚糖2.四肽及连接方式四肽中N端的Ala上α-NH2与M中乳酸的羧基连接。3.五聚Gly及连接方式(1)五聚Gly的N端α—NH2与四肽C端Ala上的羧基连接。(2)五聚Gly的C端羧基与另一个四肽的Lysε-NH2连接。溶菌酶能水解G-M间的β-1.4糖苷键,使细胞壁出现孔洞,基至解体,从而杀死细菌。人的眼泪中存在大量的溶菌酶,某些噬菌体在感染宿主时也可分泌溶菌酶。鸡蛋中也含大量的溶菌酶。生素能抑制肽聚糖的生物合成。
18一、糖脂见脂类