


- 1.40 MB
- 14页


- 1、本文档共5页,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,可选择认领,认领后既往收益都归您。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细先通过免费阅读内容等途径辨别内容交易风险。如存在严重挂羊头卖狗肉之情形,可联系本站下载客服投诉处理。
- 文档侵权举报电话:19940600175。
第二章配子发生与受精1、精子发生的一般过程:原始生殖细胞,精原细胞增殖期,初级精母细胞生长期,成熟分裂期,精子形成期2、3、精卵发生区别:1)初级卵母细胞的分裂不均等:产生含有所有细胞质的卵母细胞和不含胞质的第一极体2)卵母细胞具有丰富的内含物4、卵泡生长:原始卵泡周期性进入生长期,使卵母细胞增大,颗粒细胞增多,只有少数与促性腺激素分泌周期相同的卵泡才能存活二、受精(fertilization)受精是两性细胞融合并创造出具备源自双亲遗传潜能的新个体的过程卵母细胞成熟->精子获能->精卵间接触和识别->精子入卵->卵的激活并开始发育1、精子获能(capacitation):是指射出的精子在若干生殖道获能因子作用下,精子膜发生一系列变化,进而产生生化和运动方式的改变。意义:使精子准备顶体反应;促使精子超活化,以便通过透明带。2、精卵识别:距离识别:常见体外受精的水生生物接触识别:常见体内受精的哺乳动物(1)精子的向化性:卵子释放的精子激活肽(2)精子表面蛋白:半乳糖基转移酶,透明带附着分子(SP56),P95分子(3)参与配子间质膜相互作用的一些具粘附作用的分子:Fertilin,Cyritestin,Integrinetal.(4)顶体反应的调控机制(离子调控,脂质调控,磷酸肌醇调控)3、配子遗传物质融合:雌雄原核融合;雌雄原核的不均等性(哺乳动物)遗传印记现象;卵质重排;卵裂准备4、卵子激活1)快速阻止多精受精:电势改变产生的快速阻止:第一个精子与卵质膜结合后的1-3秒内,因钠离子流入导致膜电位迅速升高,阻止其他精子的结合2)多精受精的慢速抑制:形成受精膜的慢速阻止:受精后20-60秒内,质膜下的皮质颗粒与质膜融合,释放内含物形成受精膜,阻止其他精子进入
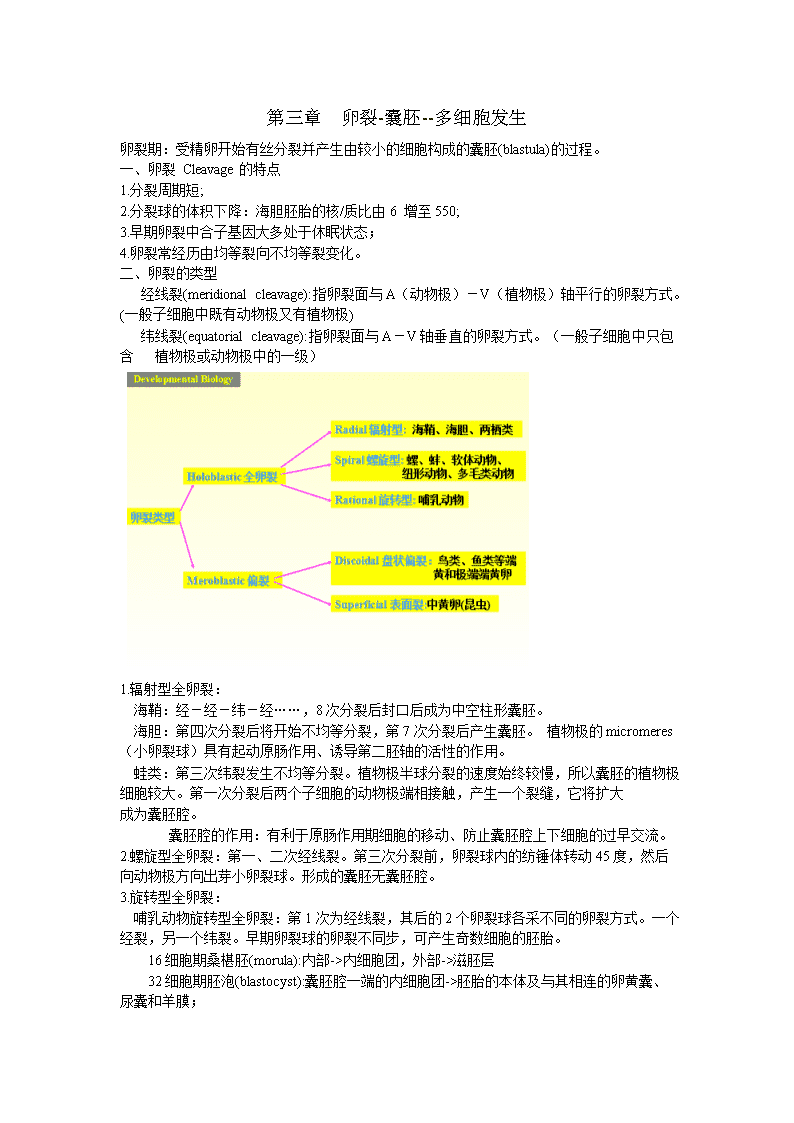
第三章卵裂-囊胚--多细胞发生卵裂期:受精卵开始有丝分裂并产生由较小的细胞构成的囊胚(blastula)的过程。一、卵裂Cleavage的特点1.分裂周期短;2.分裂球的体积下降:海胆胚胎的核/质比由6增至550;3.早期卵裂中合子基因大多处于休眠状态;4.卵裂常经历由均等裂向不均等裂变化。二、卵裂的类型经线裂(meridionalcleavage):指卵裂面与A(动物极)-V(植物极)轴平行的卵裂方式。(一般子细胞中既有动物极又有植物极)纬线裂(equatorialcleavage):指卵裂面与A-V轴垂直的卵裂方式。(一般子细胞中只包含植物极或动物极中的一级)1.辐射型全卵裂:海鞘:经-经-纬-经……,8次分裂后封口后成为中空柱形囊胚。海胆:第四次分裂后将开始不均等分裂,第7次分裂后产生囊胚。植物极的micromeres(小卵裂球)具有起动原肠作用、诱导第二胚轴的活性的作用。蛙类:第三次纬裂发生不均等分裂。植物极半球分裂的速度始终较慢,所以囊胚的植物极细胞较大。第一次分裂后两个子细胞的动物极端相接触,产生一个裂缝,它将扩大成为囊胚腔。囊胚腔的作用:有利于原肠作用期细胞的移动、防止囊胚腔上下细胞的过早交流。2.螺旋型全卵裂:第一、二次经线裂。第三次分裂前,卵裂球内的纺锤体转动45度,然后向动物极方向出芽小卵裂球。形成的囊胚无囊胚腔。3.旋转型全卵裂:哺乳动物旋转型全卵裂:第1次为经线裂,其后的2个卵裂球各采不同的卵裂方式。一个经裂,另一个纬裂。早期卵裂球的卵裂不同步,可产生奇数细胞的胚胎。16细胞期桑椹胚(morula):内部->内细胞团,外部->滋胚层32细胞期胚泡(blastocyst):囊胚腔一端的内细胞团->胚胎的本体及与其相连的卵黄囊、尿囊和羊膜;
外层的滋胚层->绒毛膜胚泡的着床:胚泡在向子宫移动过程中体积增大,是因为定位于Na-K泵将外部Na+泵入腔中,最后通过渗透作用吸水使囊胚腔增大。胚胎外的透明带阻止了胚泡与输卵管壁的粘连。胚泡到达子宫时,胚胎细胞分泌strypsin(一种类胰蛋白酶),它使透明带穿孔,胚泡从孔中挤出与子宫壁接触,通过一系列反应而着床。人类的同卵双生的发生:1、发生在滋胚层形成前(约受精后5天前)的分割,有独立的绒毛膜和羊膜。占同卵双生的33%。2、发生在滋胚层形成后但羊膜形成前(约受精后5-9天)的分割,共用绒毛膜,有独立的羊膜。占同卵双生的66%。3、发生在羊膜形成后(约受精9天后)的分割,共用绒毛膜和羊膜,易出现连体儿。嵌合胚:早期卵裂球有同等的发育潜力。卵裂球基因不一样导致。自然人群中也出现过同时有XX型和XY型细胞的人。4.盘状偏裂鸟类:前3次卵裂经线裂,发生在输卵管中,胚盘为单细胞层,仍与卵黄相接触。上胚层将形成胚胎本体;下胚层将产生胚外结构鱼类:前5次卵裂均为经线裂,产生的32个细胞为单层分布于卵黄上。其后的分裂方向不规则。囊胚(经历了10次卵裂)的三类细胞:1.卵黄多核层(YSL):胚盘的植物极边缘细胞裂解,其核和质与卵黄细胞融合在一起而构成的一层细胞核层。在胚盘下包中,部分YSL细胞核移向胚盘下成为internalYSL,它们可能起提供营养的作用;边缘处的为externalYSL,它们可能起驱动下包的作用。2.Envelopinglayer(EVL):位于胚盘最外层已表皮化的细胞,发育后期会脱落。3.DeepCells:介于YSL和EVL之间的细胞,它们将发育为胚胎本体。晚期囊胚的深层细胞的命运已经建立。5.表面裂果蝇:受精卵的合子核位于卵黄中央,胞质已被挤在卵黄与细胞膜之间成为卵周质。前13次分裂仅为细胞核的分裂,这期间的胚叫多核胚(syncytialblastoderm)。在第14次分裂时,卵膜内陷,将每个核围成一个细胞,形成细胞化胚。早期卵裂的细胞周期1、早期卵裂细胞的分裂速度快,细胞周期时相只有S,M期2、卵裂细胞特殊性的来源:卵细胞质中蛋白质、mRNA和其他营养物质的储备、原肠作用原肠作用:囊胚细胞有规则的移动,使细胞重新排列,用来形成内胚层和中胚层器官的细胞迁入胚胎内部,而要形成外胚层的细胞铺展在胚胎表面。原肠作用期的胚胎叫原肠胚主要细胞迁移类型:外包(epiboly):表皮层做为一个整体扩展,使胚胎的内层被覆盖。内陷(invagination):指胚胎局部区域的内陷。内卷(involution):指正在扩展的外层向内卷折,并沿外层细胞内表面扩展。内移(ingression):指表层的单个细胞迁入胚胎的内部。分层(delamination):指一个细胞层分成两层或多层平行的细胞层。
会聚伸展(convergentextension):指细胞间相互插入,使所在组织变窄、变薄,并推动组织一定方向移动。2、表皮细胞:细胞与细胞间紧密连接成管状或片层状结构,局部或整个结构一起运动。间质细胞:细胞与细胞间松散相连,每个细胞为一个行动单元1、海胆原肠作用的机制:内胚层的早期内陷机制:纤丝收缩使细胞变位契形,成为细胞内陷的原动力。内胚层的晚期内陷机制:伪足的收缩和细胞间的变形重排(会聚伸展)。海胆原肠作用过程:原植物极中央细胞内陷进入囊胚腔,表皮细胞转变成为初级间质细胞,然后内胚层表皮细胞内陷和扩展,其前端表皮细胞转化为次级间质细胞。两种间质细胞都将长出伪足,后者起定向和驱动细胞移动的作用。初级间质细胞在中胚层和内胚层相交处形成一圈,在腹侧有一分支延伸,它们将用于骨的形成。小分裂球启动原肠作用,可诱导第二胚轴的形成。2、爪蟾的原肠作用原肠作用开始于瓶状细胞的形成和内突。它们的内突使囊胚表面形成一个小沟,即为胚孔3、斑马鱼的原肠作用原肠作用开始于胚盘细胞向植物极方向的卵黄下包,由YSL驱动,使胚盘变薄。胚层的形成:50%下包时,与卵黄交界处的deepcells内卷,使交界处形成厚实的一圈,叫germring。内卷的细胞和由上层内移的细胞形成下胚层、上层的deepcells为上胚层。胚盾:因深层细胞的内卷和会聚扩展而在germring的某处形成的加厚区。它为胚胎的背部,从此处内卷的细胞将与其它会聚扩展的下胚层细胞一起沿背部中线形成脊索中胚层下胚层细胞还将生成近轴中胚层和内胚层。4、鸡胚的原肠作用原条的形成:由后部边缘区的上胚层细胞加厚而成,它的出现确定了胚胎的A-P轴线。原条内会形成一个凹陷,叫原沟,是上胚层细胞进入囊胚腔的门户。原条的头部末端是一个加厚层,叫Hensen`snode,是一个诱导中心。5、哺乳动物的原肠作用上胚层和下胚层的形成:在原肠作用开始时,内细胞团分裂为两层。与囊胚腔接触的一层为下胚层,将用于形成yolksac;另一层为上胚层。上胚层细胞间的缝隙将合并、扩大成为羊膜腔,腔中的液体可防止胚胎脱水和保护胚胎受振荡。上胚层将发育为胚胎的本体第五章三胚层-器官发生一、外胚层与中枢神经系统(一)中枢系统的形成神经管(neuraltube):中枢神经系统的原基Neurulation:神经管的形成外胚层细胞的命运:背部中线区的细胞将形成脑和脊髓;中线区外侧的细胞将生成皮肤;上述二者相交处的细胞为神经嵴细胞(neuralcrest),它们将迁移各处形成外周神经元、色素细胞、神经胶质细胞等。神经管形成的起始:来自背部中胚层的信号诱导预置神经板边缘的细胞的背侧收缩,而预置的表皮细胞向中线移动,使表皮与神经板交接处凸起形成神经褶。
1.Primaryneurulation:由外胚层细胞增殖、内陷并最终离开外胚层表面而形成中空的神经管。绝大多数脊椎动物前部神经管的形成采用此种方式。2.SecondaryNeurulation神经管由胚胎内细胞组成的实心索中空而成。鸟类、哺乳类、两栖类动物胚胎的后部神经管及鱼类胚胎的全部神经管的形成采取此种方式。Ø神经嵴细胞→发生部位:神经管闭合处的神经管细胞和与神经管相接的外表层细胞,它们间质细胞化而成为神经嵴细胞。→特点:具有迁移性。→分化命运:因发生的部位和迁移目的地不同而不同。可分化为感觉、交感及副交感神经系统的神经元和胶质细胞;肾上腺髓质细胞;表皮中的色素细胞;头骨软骨和结缔组织等。(二)表皮及其衍生物的发育1表皮细胞的起源胎皮和基底层基底层-棘层-颗粒层-过渡型细胞-角质层二、中胚层神经胚期中胚层分为5个区1位于胚胎背面中央的脊索中胚层—脊索2背部体臂中胚层—体节和神经管两侧的中胚层-背部许多结缔组织3中段中胚层—泌尿系统和生殖器官4离脊索稍远的侧板中胚层—心脏、血管、血细胞等5头部间质—面部组织和肌肉三、内胚层咽、消化器官、呼吸器官第六章细胞分化和胚胎诱导一、细胞分化细胞分化:同群结构与功能相同的细胞发生一系列的内外变化,成为结构与功能不同细胞的过程。(二)特点:
1)基因表达上的变化,导致组织特异性蛋白的产生;2)不同细胞在蛋白质组成上的差异导致细胞结构的不同,改变其组成就可改变其形状;3)在细胞分化的早期,不同细胞间的差异难以检测;4)分化是渐进过程,进入终端分化的细胞往往不再分裂,而终端分化后能够继续分裂的细胞可以维持和传递终端分化状态;5)细胞分化由许多细胞外信号(如细胞表面蛋白、分泌蛋白)控制。(二)细胞分化的分子机制1、基因活性状态的可逆性2、DNA的不可逆改变导致的细胞分化:体细胞重排3、基因活性状态的维持和传递决定于调控蛋白和DNA的修饰4、细胞分化中基因的转录调控:1)基因转录调控涉及通用转录因子和组织特异性转录因子2)外源信号对基因转录的激活作用5、细胞分化中RNA加工水平上的调控:RNA剪接6、细胞分化中翻译水平上的调控1)mRNA寿命的调控2)激素促进转录和增加mRNA寿命7、细胞分化中翻译后水平上的调控(三)卵母细胞中mRNA的翻译调控机制1、mRNAmasking:mRNA与其它蛋白结合成ribonucleoprotein(RNP)complex,阻止与ribosome结合;卵成熟或受精后,离子强度改变或蛋白磷酸化等导致RNP解体,翻译得以进行。2、5’-Cap的调控:如某些种类的moths(娥),其卵中的部分mRNA的5`-鸟苷酸在受精后才甲基化,然后开始翻译。3、mRNAsequester(隔离):指mRNA被阻隔于蛋白合成装置。如海胆未受精卵的histonemRNA定位于核中,受精后核破裂,mRNA才能进入胞质开始翻译。4、翻译效率的调控:如将海胆卵母细胞裂解液的pH从自然状态下的pH6.9提高到(受精后自然状态下的)pH7.4,蛋白质合成量急剧增加。受精后pH升高的作用可能包括去除mRNA的封闭蛋白和激活翻译起始因子。5、Poly(A)对翻译的调控:在小鼠的未成熟卵母细胞质中可以翻译的mRNA具有较长的poly(A),减数成熟分裂后poly(A)降解,翻译终止。在减数成熟分裂前不表达的mRNA的poly(A)较短(15-90A)。减数成熟分裂后这些mRNA迅速加上一个长的polyA,开始翻译。二、胞质定域胞质定域:细胞质物质在卵细胞质中呈一定形式分布,受精时发生运动,被分隔到一定区域,并在卵裂时分配到特定的卵裂球中,决定卵裂球的发育命运。如生殖细胞决定子三、胚胎诱导胚胎诱导:在胚胎发育过程中,相邻细胞或组织间通过相互作用,决定其中一方或双方细胞的分化方向。1、初期胚胎诱导:在原肠形成时脊索中胚层诱导其表面覆盖的外胚层形成神经板的现象诱导机理:归因于一些可溶性因子,这些因子是由母源性RNA所编码的,在早期胚胎中,他们由植物半球的细胞产生。这些因子被认为是通过胞吐作用释放进细胞间隙。2、次级胚胎诱导和三级胚胎诱导
3、感受性:胚胎组织者具有接受诱导刺激的反应能力反应组织:有感受性的组织初级感受性:尚未决定的外胚层所具有的感受性,具有时间性;次级感受性:已经决定了的组织对诱导刺激的感受性。感受性的特性:时间模式;区域模式;种间差异;遗传因子4、诱导组织的相互作用类型A、指导互作:需要从诱导细胞发出一个信号,才能启动反应细胞新基因的表达,没有诱导细胞,反应细胞就不能按特定的方式分化。B、容许互作:反应组织包含所有需要表达的潜力,只是需要允许这些特征表达的环境,并不改变所产生细胞的类型,而仅仅是让它有能力表达。5、胚胎细胞定型的两种方式是什么?1)镶嵌型发育的胚胎细胞定型:又细胞内的形态决定发生自决定,称为自主特化,整体胚胎像是自我分化的各部分组装在一起的集合体。2)调整型发育的胚胎细胞定型:由细胞相互关系来决定,称为有条件特化或渐进式特化或依赖式特化。第七章胚轴形成--果蝇胚轴形成的分子基础1、bicoid:在未受精卵中,bicoidmRNA定位在胞质前端;其受精后翻译出的蛋白质浓度沿AP轴依次降低。2、Nanos:决定后部区的发育,浓度梯度沿AP轴升高,其作用是与hunchbackmRNA结合,阻止后者在后区的翻译,帮助形成Hunchback蛋白梯度。3、Hunchback:母体mRNA在卵中均匀分布,受精后前区高浓度的Bicoid蛋白激活合子hunchback基因的表达,从而帮助形成hunchback蛋白浓度梯度。4、末端系统—Torso信号途径:未受精前,torso已均匀地分布在卵的质膜上。但其配体torsolike定位在两端的卵外膜上,不能与torso结合。受精时,torsolike得以释放,torsolike与torso结合,torso活化,启动信号传导。一、AP轴极性确定:前:bicoid从前到后浓度降低,前区高浓度的Bicoid蛋白激活合子hunchback基因的表达,从而形成hunchback蛋白从前到后降低的浓度梯度,决定头胸的形成。后:nanos从后到末浓度升高,与hunchbackmRNA结合,阻止后者在后区的翻译,形成Hunchback蛋白从后到末降低的浓度梯度,决定腹部的形成。末:未受精前,torso已均匀地分布在卵的质膜上。但其配体torsolike定位在两端的卵外膜上,不能与torso结合。受精时,torsolike得以释放,torsolike与torso结合,torso活化,启动信号传导,决定原头原尾的形成。Hunchback,torso为其它Gap基因的表达提供位置信息,导致AP轴线区域化二、DV轴极性确定:在受精前腹部的卵泡细胞合成Pipe、Windbeutel、Nudel,定位在腹部卵外膜上,受精后经一系列反应,最终使卵周质腹部区的Dorsal蛋白释放进入核中,启动腹部特异性基因的表达。三、体节:1、副体节由pair-rule基因活性界定副体节:在原肠作用开始后,胚胎表面沿AP轴线出现一些过渡性的浅沟,将胚胎分为14个区域。
体节极性基因:在pair-rule基因表达之后立即表达的基因,它们决定了体节的边界和体节内细胞的命运。Engrailed是确定副体节和体节边界的关键基因,其表达受高浓度fushitarazu(ftz)和even-skipped(eve)的激活。不同体节的发育命运决定于homeoticselectorgenes(同源异形选择基因)第八章性别决定1.Primarysexdetermination:指生殖腺发育为睾丸或卵巢的选择。胚胎生殖腺(gonad)的发育命运决定于其染色体组成,Y染色体的存在使生殖腺的体细胞发育为testis而非ovary。生殖嵴双向性腺滤泡细胞卵泡支持细胞间质细胞附睾输精管阴茎子宫近端阴道2.secondarysexdetermination:指睾丸或卵巢形成后,由它们分泌的激素来影响性器官的发育。卵泡膜细胞抗中肾旁管激素(anti-Mullerianducthormone,AMH):由睾丸支柱细胞分泌的560aa糖蛋白,其作用可能是诱导中肾旁管周围的间质细胞分泌一种促凋亡因子,使中肾旁管退化。睾丸酮(testosterone):由睾丸间质细胞合成,其作用是诱导中肾管分化为输精管、精囊、附睾。二氢睾丸酮(dihytestosterone):由睾丸酮转变而成,其作用是控制外生殖器官的形成。2、性别异常 1)男性假两性畸形核型为46,XY;X染色质检查为阴性,Y染色质检查为阳性,但呈女性表型。①XY性腺退化症46,XY②睾丸女性化综合征46,XY2)女性假两性畸形核型为46,XX;X染色质检查是阳性,Y染色质检查为阴性,但接近男性的表型。①肾上腺皮质过度发育综合症46,XX
②较罕见的45,X男性3)真两性畸形①雌雄嵌合体(chimaera)②XX真两性畸形46,XX3、性比值:在人类大群体之中,男女人数的比例总的来说基本上接近1:1的,但在不同年龄区段范围内,男女之间的性比值则常有不同的差异,一般可大致分为如下三种类型:1)受精时的性比值(第一性比值):X:Y是120:100。①Y精子比X精子小而轻,运动速度快,与卵子先接触的机会大些;②Y精子比X精子存活力稍强;③卵子表面更容易接受Y精子。2)出生时的性比值(第二性比值):X:Y是103:100-105:100男胎的死亡率显著大于女胎3)产后的性比值(第三性比值):通常是指成年人的性比值。1:1男性的死亡率比女性偏高一、果蝇的性别决定第九章发育异常与癌症1、发育异常:有机体在发育过程中,经受各种外界和内部因子的作用,这些作用可能引起畸胎瘤(teratoma)即发育异常癌症:是肿瘤的一种,是指细胞调控机制发生缺陷,并导致恶性和侵犯性肿瘤形成的一种疾病。是一种恶性肿瘤。2、癌症的特征(1)接触抑制的丧失(2)粘着性下降(3)凝聚性增强(4)产生新的膜抗原(5)无限增殖
3、癌基因和抑癌基因(1)癌基因(oncogene)是一类会引起细胞癌变的基因分类:病毒癌基因:指反转录病毒的基因组带有可使受病毒感染的宿主细胞发生癌变的基因,简写成v-onc细胞癌基因:指正常细胞基因组中,一旦发生突变或被异常激活后可使细胞发生恶性转化的基因,简写成c-onc,又称原癌基因(2)抑癌基因又称肿瘤抑制基因(tumorsuppressorgene)或抗癌基因(anti-oncogene)是指能够抑制细胞癌基因活性的一类基因,其功能是抑制细胞周期,阻止细胞数目增多以及促使细胞死亡。癌基因和抑癌基因的区别:癌基因只要有一个等位基因基因发生突变时就可以发生癌变;而抑癌基因只要有一个等位基因是野生型时,就可以抑制癌症的发生。4、致癌的可能机制癌症可能是由于生长的控制受到干扰,或者部分细胞未能从增殖转变为终末分化而引起。缺陷可以发生在控制体系的各个分子级水平上。1)生长因子2)信号的接受受到干扰3)信号传导受到干扰4)细胞核内对信号的影响受到干扰第十章衰老和死亡一、细胞衰老的一般特征:1、细胞质膜变性2、细胞器降解3、细胞核异常4、细胞水分减少二、衰老的基本学说1、自由基理论(freeadicaltheory)生命活动离不开氧,而生物氧化过程中的中间产物能够导致细胞结构和功能改变。自由基是指带有奇数电子数的化学物质,它们都带有未配对的自由电子,这些自由电子导致了这些物质的高反应活性。生物氧化、辐射、酶促反应等过程都会释放自由基。细胞内产生的自由基可以被清除或限制在某一区域,防止对细胞产生的危害。清除和限制的方式:生物体内的抗氧化分子,如维生素C、E等都能和自由基结合,终止自由基的扩增反应;利用细胞内的超氧化物歧化酶和过氧化氢酶,协同清除自由基;细胞内部形成自由基隔离,使自由基只能局限在特定部位。但细胞内过多的自由基不能被清除,对细胞造成伤害。2、细胞的程序化死亡(programmedtheory)生物体的衰老是由基因组上的基因控制的,这些基因按照发育的时空进行顺序表达,决定某些组织器官或细胞群按计划分裂、生长、分化、死亡,称细胞的程序化死亡。三、细胞程序化死亡与细胞坏死的区别:
特征程序化死亡坏死细胞形态凝聚、断裂溶解膜完整性保持到最后阶段很早消失线粒体完整肿胀染色质边缘化固缩核生化变化DNA降解,电泳梯形带DNA弥散降解,电泳拖影炎症反应周围活组织无炎症反应周围活组织有炎症反应谷氨酰胺转移酶活性升高无变化四、细胞程序化死亡的生物功能:(1)清除无用的或多余细胞;(2)除去不再起作用的细胞;(3)除去发育不正常的细胞;(4)除去一些有害的细胞。花的发生-SAMMeristerm(分生组织):指植物中的胚性组织,或一群未分化、正在生长和活跃分裂的细胞茎端分生组织shootapicalmeristem(SAM):最小的叶原基上方茎端区域腋芽分生组织(axiallybudmeristem)一、SAM分区Ø中央区:是分生组织的原始细胞,由一群未分化并能自我更新的细胞组成,细胞相对较大,分裂缓慢。中央区干细胞分裂后产生2部分细胞,一部分保留在中心区域形成干细胞后裔,始终保留在原来位置,保持分裂潜能;分裂出来的另部分叫子细胞,子细胞以较快的速度分裂,离开中央区到分生组织的周围区。Ø周缘区:由位于分生组织侧翼的一群相对较小迅速分裂的原始细胞组成,周缘区是叶原基的起始位点,发育成器官。Ø肋状分生组织:位于中央区下方,它们的分裂和伸长导致茎的延伸,是茎髓的起始位点。SAM从外到内可分为3层,L1,L2和L31.L1:无叶绿素的叶表皮2.L2:栅栏层和部分海绵组织3.L3:维管束和部分海绵组织二、持茎端分生组织正常功能的基因:1、STM(shootmeristemless):SAM形态发生强STM功能缺失突变体中,胚胎发生过程中不能形成SAM,STM是分生组织建立和保持所必须的。主要功能:抑制分生组织中细胞的分化,保证分生组织内细胞的扩增,从而能有足够数量的细胞成为侧生器官原基。2、WUS(Wuschel):保持分生细胞在一定的数量水平。变体不像正常的茎尖分生组织呈现圆顶形,而是又扁又薄。然而,在茎的其他位置有
产生新的叶原基和二级茎分生组织,产生成百上千的叶子。3、CLV(Clavata):维持周缘分生区和茎尖分生组织细胞数量的平衡。clv突变体丧失维持茎尖分生组织细胞数量平衡的功能,CLV1,CLV3是限制中心区大小所必须的Transitiontoflowering1、无限花序:其花序分生组织能不断分化出花原基,在花序轴不断生长延长的同时,在花序轴侧部不断形成次级花序枝或小花,花序主轴的顶端不被决定成一朵顶花序。2、有限花序:花序分生组织只能分化出一定数目的花原基,然后花序主轴的顶端被决定转变成花分生组织从而形成一朵顶花,停止分化。主轴的顶花一般最早被决定。花序中最顶点或最中的花先开,由于顶花的开放,限制了花序轴顶端继续生长,因而以后开花顺序渐及下边或周围、Floraldevelopment
拟南芥中有3个花分生组织特异基因:花序原基到花原基的转变需要一些基因的表达,包括AP1,CAL和LFY基因Ap1和lfy突变体最终会产生花的结构,因为AP1和LFY功能可以部分补偿功能的缺失。Ap1lfy双突变体中,花完全转化为茎SUP基因是模式植物拟南芥控制花器官调控基因之一,它能抑制B类功能基因在雌蕊中的表达Cal-1突变体基本上看不到表型上的变化在ap1和Cal-1突变体中,花分生组织的形成过程被阻断,因此突变体的花看起来像花椰菜的顶端组织。同源异型突变(homeoticmutation):有时花某一重要器官位置发生了被另一器官替代的突变,如花瓣部位被雄蕊替代,这种遗传变异现象称为花发育的同源异型突变。同源异型基因(homeoticgene):控制同源异型化的基因称为同源异型基因。
第1轮处于A功能控制之下,原基发育为萼片。第2轮受A和B两个功能的控制,原基发育为花辩。第3轮由B和C两个功能控制,原基发育为雄蕊。第4轮则由C功能控制,原基发育为心皮。花瓣雄蕊心皮萼片ABCDE模型胚珠